Human liver development begins during the third week of gestation and does not achieve mature architecture until about 15 years of age. It reaches its largest relative size, 10% of fetal weight, around the ninth week. It is about 5% of body weight in the healthy neonate. The liver is about 2% of body weight in the adult. It weighs around 1400g in an adult female and about 1800g in the male.*
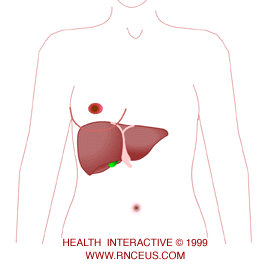
The liver is located in the right upper quadrant
of the abdomen, just below the diaphragm. It is almost completely behind the rib cage but the lower edge may be palpated along the right costal margin during inspiration. A connective tissue
layer called Glisson's capsule covers the surface of the liver. The capsule extends to invest all but the smallest the vessels within the liver. * The falciform ligament attaches the liver to the abdominal wall and diaphragm and divides the liver into a larger right lobe and a smaller left lobe.
In 1957, the french surgeon Claude Couinaud described 8 liver segments. Since then, radiographic studies describe an average of twenty segments based on distribution of blood supply*. Each segment has its own independent vascular and biliary branches. Surgeons utilize these independent segments when performing liver resection for tumor or transplantation. There are at least three reasons why segmental resection is superior to simple wedge resection. First, segmental resection minimizes blood loss because vascular density is reduced at the borders between segments. Second, it results in improved tumor removal for those cancers which are disseminated via intrasegmental branches of the portal vein. Third, segmental resection spares normal liver allowing for repeat partial hepatectomy*.
Each segment of the liver is further divided into lobules. Lobules are usually represented as discrete hexagonal aggregations of hepatocytes. The hepatocytes assemble as plates which radiate from a central vein. Lobules are served by arterial, venous and biliary vessels at their periphery. This model is useful for teaching purposes but more closely resembles the adult pig lobule than the human. Human lobules have little connective tissue separating one lobule from another. The paucity of connective tissue makes it more difficult to identify the portal triads and the boundaries of individual lobules. Central veins are easier to identify due to their large lumen and because they lack connective tissue that invests the portal triad vessels.
Lobules consist of hepatocytes and the spaces between them. Sinusoids are the spaces between the plates of hepatocytes. Sinusoids receive blood from the portal triads. About 25% of total cardiac output enters the sinusoids via terminal portal and arterial vessels. Seventy-five percent of the blood flowing into the liver comes
through the portal vein; the remaining 25% is oxygenated blood
that is carried by the hepatic artery. The blood mixes, passes through the sinusoids, bathes the hepatocytes and drains into the central vein.
About 1.5 liters of blood exit the liver every minute.
The liver is central to a multitude of physiologic
functions, including:
Clearance of damaged red blood cells & bacteria by phagocytosis
Nutrient management
Synthesis of plasma proteins such as
albumin, globulin, protein C, insulin-like growth factor, clotting factors, etc.
Biotransformation of
toxins, hormones, and drugs
Vitamin & mineral storage
Phagocytosis
Red blood cell (RBC) life span is about 120 days. Reticuloendothelial (macrophage) cells in the spleen, liver and bone marrow are primarily responsible for clearing pathogens and debris. Kupffer cells are reticuloendothelial cells resident in the liver sinusoids that scavenge damaged RBCs and bacteria as they pass through. Hundreds of millions of RBCs are removed by the reticuloendothelial system every minute. Kupffer cells, like other reticuloendothelial macrophages, lyse RBCs into heme and globin. Globin is further catabolized into polypeptide components for reuse. Heme is broken into biliverdin and iron. Biliverdin is converted to bilirubin. Iron is transported by transferrin to the liver and spleen for storage and to the bone for hematopoiesis.
About 85% of bilirubin is derived from lysis of RBCs, the rest comes from the breakdown of other hemoproteins like myoglobin, cytochromes and peroxidases. Kupffer cells like other reticuloendothelial macrophages release bilirubin into the blood. In the blood, bilirubin binds to albumin. The albumin/bilirubin compound is small enough to pass through the endothelial fenestrae and into the space of Disse where it contacts the hepatocyte.
Hepatocytes cleave bilirubin from albumin and absorb the bilirubin. In the hepatocyte cytoplasm, bilirubin is conjugated to glucouronic acid. Bilirubin uridine diphosphate glucuronyl transferase (UDPGT) catalyzes the bonding of glucuronic acid and bilirubin to produce water-soluble bilirubin. Water soluble conjugated bilirubin is secreted into canaliculi along with water, electrolytes, bicarbonate, bile acids, salts, cholesterol and phospholipids. This combination is called bile and serves as a detergent to keep bile soluble in the biliary tract. Bile drains from the canaliculi>canal of Hering>bile ducts>common hepatic duct>gallbladder>common bile duct>ampulla of vater>duodenum.*
In the duodenum, bile salts attach to fat globules forming smaller micelles that collect fatty acids and glycerol. The micelles travel to the jejunum where they deliver their cargo to the intestinal epithelium. Inside the epithelial cells glycerol and fatty acids are rejoined to form triglycerides. Finally triglycerides are joined to cholesterol and proteins are added to the surface; creating a chylomicron.
The liver receives a variety of lipid forms including: chylomicrons remnants, very low density lipoproteins (VLDL), low density lipoproteins (LDL), high density lipoproteins (HDL) and fatty acids. Large lipoprotein molecules are broken into smaller units by the lytic action of lipoprotein lipase (LPL) expressed on endothelium of vessels. Circulating lipoproteins small enough to enter the space of Disse attach to receptors on the hepatocyte. These lipoprotein remnants are held near the heptocyte surface and exposed to hepatic lipase compounds. Low Density Lipoprotein receptors transfer the lipoprotein fragments into the hepatocyte by the process of endocytosis.
Chylomicrons are the product of intestinal packaging of dietary fats. Chylomicrons are produced in the duodenal villi and secreted into the lymph lacteals for delivery to the thoracic duct>subclavian vein>superior vena cava>right ventricle>lungs>Left ventricle>aorta>hepatic artery>sinusoid. Chylomicrons range from 75-1200nm in diameter. They contain 98% lipids and 2% protein. Chylomicrons are degraded in the blood by contact with LPL. Chylomicrons become smaller and more dense as fatty acids are stripped off. Loss of fatty acids results in chylomicron remnants of various sizes and density when they finally reach the liver. Hepatic lipase expressed by the hepatic sinusoidal endothelium and hepatocytes continues the remnant degradation.
Very low density lipoproteins (VLDL) are synthesized primarily in the hepatocyte. VLDLs range from 30-80nm. They contain 90% lipids and 10% protein.Their purpose is to transport triglycerides made in the liver into plasma for use or storage outside the liver.
Low density lipoprotein (LDL) is formed from VLDLs in the plasma by the action of lipase. LDL diameter is about 20nm. They contain 70% lipids and 30% protein. LDLs distribute cholesterol throughout the body. Cholesterol is an important constituent of: VLDL, cell membranes, hormones, bile etc.
High density lipoprotein (HDL) are small lipoprotein particles (5-15nm) formed in the liver and intestine. They range from 5-15nm in diameter. They contain 50% lipids and 50% protein. HDLs collect cholesterol & lipoprotein fragments from the blood and blood vessel plaques and return them to the liver for repurposing.
Fatty acids are linear hydrocarbon chains that are the major constituents of dietary lipoproteins (triglycerides). The liver degrades lipoproteins with hepatic lipase or synthesizes fatty acids from carbohydrate sources. When carbohydrate energy sources are low fatty acids are oxidized for energy.
Carbohydrate management:
When energy intake exceeds energy output the liver stores the surplus glucose as glycogen. When energy output exceeds energy intake the liver reacts by releasing stored energy as glucose.
Glucose is the preferred energy source for most tissues but the body maintains very limited supplies of free circulating glucose. Certain tissues like the brain, RBCs, lens and cornea use glucose almost exclusively. To supply these tissues when blood glucose drops the liver lyses glycogen. Glycogen is a complex molecule composed of thousands of glucose units. Hepatocytes and myocytes store glucose as intracellular glycogen granules.
The liver is central to blood glucose management because the liver is the only organ that can store and release glucose into the blood for use by other organs. After a meal the liver removes excess blood glucose and stores up to 8% of its weight as glycogen. Myocytes can store about 1-2% of total muscle mass as glycogen but once glucose enters a myocyte it must be used or stored by that myocyte. Myocytes lack the enzyme necessary to release glucose into the circulation.
The liver uses three metabolic processes to manage carbohydrates and insure adequate blood glucose:
Glycogenesis - excess glucose, fructose, and galactose are
converted to glycogen and stored in the liver.
Glycogenolysis - when blood glucose falls, the
liver breaks down stored glycogen to raise blood
glucose levels
Gluconeogenesis - the liver can synthesize glucose from lactic acid, some amino acids and glycerol. When glucose is low the liver can derive energy from the metabolism of fatty acids which can conserve available glucose.
Protein management:
Dietary protein is denatured by stomach acids and digested into amino acids in the small intestine. Amino acids are absorbed by the small intestine and delivered to the liver via the portal circulation. Up to 50% of the livers' energy requirements can be supplied by amino acid oxidation. Oxidative deamination breaks amino acids into keto acid and an ammonia molecule. The keto acid is used in the Kreb's cycle to produce ATP. The liver combines ammonia with CO2 to form urea and H2O.
The liver also uses dietary amino acids and those released during normal tissue catabolism to synthesize its own proteins and enzymes as well as plasma proteins. Plasma proteins produced by hepatocytes include: albumin, fibrinogen, prothrombin, a-fetoprotein, a2-macroglobin, hemopexin, transferrin, complenent components C3,C6 andC1, a1-antitrypsin, caeruloplasmin, etc.•
Albumin is only produced by the liver and equals about 50% its total protein synthesis. About 12 grams of albumin are synthesized by the normal liver daily.* Patients with decompensated cirrhosis produce only about 4 g per day. About 40% of total albumin is intrascular.
a-fetoprotein peaks about 16 weeks gestation and disappears a few weeks after birth. It may reappear in association with chronic hepatitis and a number of carcinomas
a1-antitrypsin deficiency is inherited
a2-macroglobin functions as a protease inhibitor. It is active in the inhibition of thrombin and plasmin.
hemopexin transports heme in the plasma protecting tissues from the oxidative actions of heme.
transferrin is globulin that transports heme to bone marrow for incorporation into erythroid precursors.
complement components assist the immune system to raise an immune response.
caeruloplasmin is the major copper carrying plasma protein.
Detoxification and Drug Metabolism
Hepatocytes help the body eliminate xenobiotics (toxins) by transforming them into more excretable polar compounds through a two step process. In PHASE I (Biotransformation), cytochrome p450 liver enzymes and NADPH-cytochrome c reductase catalyse hydrolysis, reduction and hydroxylation of the xenobiotic. These chemical reactions add or expose functional groups such as -OH, -SH, -NH2 or –COOH on the substance. This prepares it for conjugation with a polar, water soluable substance. PHASE II (Conjugation) liver enzymes such as Glucuronosyltransferase, Sulfotransferase, Glutathione-S-transferase and Acetyltransferase add glucuronide, sulfate, glutathione or acetyl adducts to the functional group making the conjugate more water soluble and more easily excreted in the urine and feces.
In general, Phase I and Phase II reactions serve to minimize the accumulation of foreign substances by increasing their water solubility and excretion. However, these reactions may also transform some xenobiotics from a non-toxic substance to a toxic metabolite. Acetaminophen is an example of the biotransformation of a non-toxic drug to the hepatotoxic metabolite N-acetyl-p-benzoquinoneimine by cytochrome P. Production of toxic metabolites within the liver can cause hepatic injury.
Vitamins
The liver also plays an
important role in vitamin and mineral (iron & copper) storage. About 80% of the
body's vitamin A stores are concentrated in fat droplets within the stellate cells of the liver. In pathological conditions like hepatic fibrosis or liver cirrhosis the stellate cells lose vitamin A, transform into fibroblasts or myofibroblasts and begin producing large amounts of collagen and adhesive glycoproteins.* Normal vitamin A reserves are enough to prevent a deficiency for about 10 months. The liver
also contains about a year supply of B12. Vitamin D stores equal about 3-4 months. Small amounts of Vitamins E and K and Vitamin C are stored in the liver to facilitate liver functions.
 Human liver development begins during the third week of gestation and does not achieve mature architecture until about 15 years of age. It reaches its largest relative size, 10% of fetal weight, around the ninth week. It is about 5% of body weight in the healthy neonate. The liver is about 2% of body weight in the adult. It weighs around 1400g in an adult female and about 1800g in the male.*
Human liver development begins during the third week of gestation and does not achieve mature architecture until about 15 years of age. It reaches its largest relative size, 10% of fetal weight, around the ninth week. It is about 5% of body weight in the healthy neonate. The liver is about 2% of body weight in the adult. It weighs around 1400g in an adult female and about 1800g in the male.*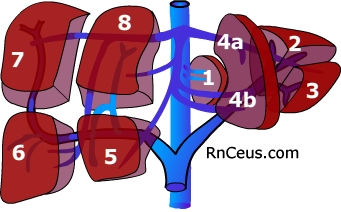 In 1957, the french surgeon Claude Couinaud described 8 liver segments. Since then, radiographic studies describe an average of twenty segments based on distribution of blood supply
In 1957, the french surgeon Claude Couinaud described 8 liver segments. Since then, radiographic studies describe an average of twenty segments based on distribution of blood supply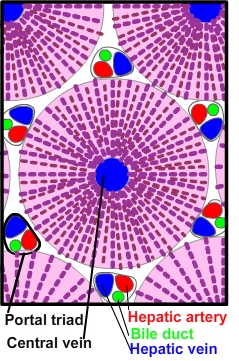 Each segment of the liver is further divided into lobules. Lobules are usually represented as discrete hexagonal aggregations of hepatocytes. The hepatocytes assemble as plates which radiate from a central vein. Lobules are served by arterial, venous and biliary vessels at their periphery. This model is useful for teaching purposes but more closely resembles the adult pig lobule than the human. Human lobules have little connective tissue separating one lobule from another. The paucity of connective tissue makes it more difficult to identify the portal triads and the boundaries of individual lobules. Central veins are easier to identify due to their large lumen and because they lack connective tissue that invests the portal triad vessels.
Each segment of the liver is further divided into lobules. Lobules are usually represented as discrete hexagonal aggregations of hepatocytes. The hepatocytes assemble as plates which radiate from a central vein. Lobules are served by arterial, venous and biliary vessels at their periphery. This model is useful for teaching purposes but more closely resembles the adult pig lobule than the human. Human lobules have little connective tissue separating one lobule from another. The paucity of connective tissue makes it more difficult to identify the portal triads and the boundaries of individual lobules. Central veins are easier to identify due to their large lumen and because they lack connective tissue that invests the portal triad vessels.
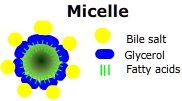 In the duodenum, bile salts attach to fat globules forming smaller micelles that collect fatty acids and glycerol. The micelles travel to the jejunum where they deliver their cargo to the intestinal epithelium. Inside the epithelial cells glycerol and fatty acids are rejoined to form triglycerides. Finally triglycerides are joined to cholesterol and proteins are added to the surface; creating a chylomicron.
In the duodenum, bile salts attach to fat globules forming smaller micelles that collect fatty acids and glycerol. The micelles travel to the jejunum where they deliver their cargo to the intestinal epithelium. Inside the epithelial cells glycerol and fatty acids are rejoined to form triglycerides. Finally triglycerides are joined to cholesterol and proteins are added to the surface; creating a chylomicron.  Fatty acids are linear hydrocarbon chains that are the major constituents of dietary lipoproteins (triglycerides). The liver degrades lipoproteins with hepatic lipase or synthesizes fatty acids from carbohydrate sources. When carbohydrate energy sources are low fatty acids are oxidized for energy.
Fatty acids are linear hydrocarbon chains that are the major constituents of dietary lipoproteins (triglycerides). The liver degrades lipoproteins with hepatic lipase or synthesizes fatty acids from carbohydrate sources. When carbohydrate energy sources are low fatty acids are oxidized for energy.